Golgi-komplex: Terminologi, Förekomst, Morfologi, Struktur och andra detaljer
Golgi-komplex: Terminologi, Förekomst, Morfologi, Struktur, Strukturell och Biokemisk Polarisering, Kemisk Sammansättning, Funktioner och Origins!
I 1898 upptäckte Golgi en retikulär struktur i cytoplasman genom en silverfärgningsmetod. Navnet "Golgi-apparat" som generellt ges till denna struktur är förvirrande eftersom det föreslår ett bestämt förhållande till cellens fysiologiska processer.
Idag verkar det lämpligare att använda namnet "Golgi substans" eller "Golgi-komplex" för att hänvisa till detta material som har speciella färgningsegenskaper. Eftersom dess brytningsindex liknar matrisen är Golgi-komplexet svårt att observera i levande celler. Användningen av elektronmikroskopet har åstadkommit en distinkt bild av denna komponent, och dess submikroskopiska struktur har uppenbarats.
I åratal trodde Golgi-komplexet vara en artefakt av olika fixerings- och färgprocedurer. Med andra ord trodde många forskare att strukturen som observerades under många mikroskopiprocedurer och kallas Golgi inte existerade faktiskt i levande cellen. Guilliermond (1923), Parat (1927), Walker och A lien (1921) uppstod tvivel om förekomsten av Golgi-komplexet.
Terminologi av Golgi-komplexet:
Holmgren hänvisade till Golgi-komplexet som trophospongium (Cajal kallade det som Golgi-Holmgren kanaler). Baker använde termen lipokondrier på grund av det antagna lipidinnehållet. Termen Dalton Complex gavs efter namnet på dess observatör Dalton 1952. Sjostrand föreslog termen cytomembran för Golgi-systemet.
Sosa har föreslagit följande nomenklatur för Golgi-komplexet:
1. Golgiokinesis :
Uppdelning av Golgi-apparaten under kärnvåning.
2. Golgiosomer :
Korpuskler som produceras av Golgiogenes kallas som Golgiosomer som beskrivs som Golgimaterial i ryggradslösa djur.
3. Golgiolys :
Golgi-apparatens upplösningsprocess.
4. Gogiorrhexis :
Fragmentering på Golgi-apparaten.
5. Golegiogenes :
Formation och differentiering av Golgi-kroppen under embryonisk utveckling.
6. Golgio-cytoarkitektur :
Studie av cellstrukturen i förhållande till Golgi-apparaten.
Förekomst:
Golgi-komplexet förekommer i alla celler utom de prokaryota cellerna (nämligen Mycoplasma, bakterier och blågröna alger) och eukaryota celler av vissa svampar, spermier av bryofyter och pteridofyter, celler av mogna siktrör av växter och mogen sperma och rött blod celler av djur.
Golgikomplexets morfologi:
Golgi-komplexets morfologi varierar från cell till cell beroende på vilken typ av cell de finns i. Två former av Golgi-komplex har observerats.

1. Lokaliserad form :
I polariserade celler av ryggradsdjur (som har bas och apex), är Golgi-komplexet enstaka och upptar i en fast position. Det ligger mellan kärnan och sekretoriska polen. Detta kan ses bäst i sköldkörtelceller, i exokrina celler i bukspottkörteln och tarmarnas slemhinnor.
2. Diffuserad form :
I vissa specialiserade celler av ryggradsdjur (nervceller och leverceller), i de flesta växtceller och i cellerna av ryggradslösa djur finns flera enheter av Golgi-komplexet spridda tillsammans med elementen i endoplasmatisk retikulum. Varje enhet kallas en diktyosom. I leverceller uppträder så många som 50 diktyosomer per cell och i vissa växtceller kan deras antal uppgå till hundratals.
Form :
Formen på Golgi-komplexet är ganska varierbar i olika somatiska celltyper av djur. Även i samma cell finns variationer i olika funktionella steg. Formen är emellertid konstant med varje celltyp. Det varierar i form från en kompakt massa till ett dispergerat trådformigt nätverk.
Siffra:
Antalet Golgi-staplar per cell varierar enormt, beroende på celltypen, från så få som en till hundratals. Det finns en enda stor i vissa celler medan i Paramoebas fall finns två. I Stereomyxa (en Amoeba-art) finns det många Golgi-komplex. Nervceller, leverceller och de flesta växtceller har också flera Golgi-komplex, där det finns cirka 50 i leverceller.
Vid algrisizoider ses mer än 25 000 Golgi-komplex (Sievers 1965). Golgi-komplexet kan till och med stå för en stor del av cellvolymen i vissa specialiserade celler. Ett exempel är tarmcellen i tarmepitelet, vilket utsöndrar slem i tarmen; glykoproteinerna i slem glykosyleras huvudsakligen i Golgi-komplexet.
Storlek:
Storleken är likaså variabel. Det är stort i nerv- och körtelcellerna och små i muskelcellerna. Generellt är Golgi-komplexet välutvecklat medan cellen är i aktivt tillstånd. När cellen växer gammal minskar komplexet gradvis i storlek och försvinner.
Placera:
Placeringen av Golgi-komplexet är relativt fast för varje celltyp. I cellerna med ektopoderalt ursprung polariseras Golgi-komplexet från tiden för embryonal tillstånd mellan kärnan och periferin (Cajal, 1914). I de sekretoriska exokrina cellerna som i allmänhet är en typisk polarisering finns Golgi-komplexet mellan kärnan och sekretoriska polen.
I endokrina körtlar är polariteten hos denna organoid variabel, förutom i sköldkörteln, där den är orienterad mot follikelns centrum. I de yngre cellerna och ofta i de äldre ligger det oftast på ena sidan av kärnan, men i vissa fall kan det helt omsluta det. I muskel-ganglioniska celler är positionen perinuclear.
Detaljerad struktur av Golgi-komplexet:
Dalton och Felix (1954) beskrev Golgi-komplexet i råtta-epididymis efter att ha tagit de första elektronmikrograferna.
Följande beskrivning av Golgi-komplexet är en sammansatt baserad på flera författares arbete:
1. Cisternae :
Cisternae eller saccules liknar den släta ytan ER, och förekommer i sektion som staplar av nära avskiljda membranavgränsade säckar. Antalet sacculer varierar från 4 till 8 i de flesta djur- och växtcellstyper. I Euglena kan antalet gå upp till 20.
Scculernas membran är ungefär 60 till 70 A i tjocklek som omsluter en hålighet ca 150 A bred, vars kanter ofta dilateras. Enligt de flesta författare finns det två väldefinierade ansikten i cisternae, dvs konvexa och konkava; den senare betecknas generellt som den mogna eller formande eller distala ytan och den konvexa sidan antas vara den omogna eller exciterade eller proximala ytan, varvid cisteranlängden i parallellmatrisen är separerad från varandra med ett utrymme av ca 200 till 300 A 0 .
Vad som håller dem tillsammans är ännu inte känt, men i få celler ses ett tunt lager elektroniskt ogenomskinligt, ibland tätt material mellan sacculerna, som i vissa regioner är mer framträdande som Amos och Grimstone (1968) tillämpade termen noder. Mollenhauer et al., (1973) undersökte i viss detalj mellanliggande element och plack i viss växt Golgi-komplex.

2. Tubuler :
Ur det yttre området av cisternae uppstår ett komplext anastomoserat platt nätverk av tubuler med en diameter av 300 till 500 A °. Clowner och Juniper (1969) har jämfört detta rörformiga nät till skivan av spets.
3. Vesiklar :
Vesiklarna är små droppliknande säckar som förblir fästa vid tubulär vid periferin av cisternae.
De har följande två typer:
(a) Smidiga vesiklar :
De släta vesiklarna har en diameter av 20 till 80 μm. De innehåller sekretoriskt material (så kallade sekretoriska vesiklar) och är böjda från ändarna av cisternal tubules i nätet. Ofta mer än en tubuleanslutning, och antagligen fyller, en enda formande vesikel.
(b) belagda blåsor :
De belagda blåsorna är sfäriska utskjutningar, ca 50 μm i diameter och med en grov yta. De finns i periferin av organellen, vanligtvis vid ändarna av enstaka tubuler och är morfologiskt ganska skilda från de sekretoriska vesiklarna. Deras funktion är okänd.
4. Golgiska vakuoler :
Dessa är stora rundade säckar som finns på Golgi. Dessa bildas antingen av den expanderade cisternae eller genom fusion av sekretoriska vesiklar. Vakuolerna är fyllda med någon amorf eller granulär substans.
Golgi-komplexet är strukturellt och biokemiskt polariserat:
Golgi-komplexet har två separata ansikten: en cis, eller formande ansikte och ett trans, eller modningande ansikte. Cis-ansiktet är nära associerat med en jämn övergångsdel av den grova ER. I sekretoriska celler är trans-ansiktet ansiktet närmast plasmamembranet: här återfinns de stora sekretoriska vesiklarna uteslutande i samband med transfasen av en Golgi-stack och membranet av en bildande sekretorisk vesikel är ofta kontinuerlig med den av Trans-ansiktet på den sista ("trans-most") cisterna.
I motsats till detta är de små Golgi-vesiklarna lokaliserade jämnare längs stapeln. Proteiner anses vanligtvis komma in i en Golgi-stapel från ER på cis-sidan och att gå ut för flera destinationer på trans-sidan; Men varken deras exakta väg genom Golgi-komplexet eller hur de reser från cisterna till cisterna längs varje stapel är kända.
Golgi-komplexets två ansikten är biokemiskt distinkta. Exempelvis kan en variation i Golgi-membrans tjocklek detekteras över stapeln i vissa fall, med de vid cis-sidan är tunnare (ER-liknande) och de vid transsidan är tjockare (plasmamembran som).
Mer slående är de resultat som erhållits när vissa histokemiska tester används i samband med elektronmikroskopi för att lokalisera specifika proteiner inom Golgi-komplexet. Några av dessa test avslöjar membranbundet enzym, aktiviteter som visar en distinkt polaritet i lokaliseringen inom Golgi-stacken.
En speciellt intressant biokemisk upptäckt var upptäckten att lysosomala enzymer, såsom sur fosfatas, är koncentrerade med de mest transiterande cisternerna i Golgi-stacken och inom några av de belagda blåsorna i närheten. Detta föreslår att specifika vesiklar som lämnar för lysosomer samlas i denna region.
Sekretoriska proteiner återfinns genom histokemiska metoder i alla de staplade cisternae, även om de stora sekretoriska blöjorna, i vilka dessa produkter är koncentrerade och endast associerade med den mest överlägsna Golgi-cisterna.
Kemisk sammansättning:
När det gäller Golgi-komplexets kemiska sammansättning har det visat sig att följande ämnen är närvarande:
1. Fosfolipider :
Fosfolipidsammansättning av Golgi-membran är mellanliggande mellan de hos endoplasmiska membran och plasmamembran.
2. Proteiner och enzymer :
Golgi-komplexet från olika växt- och djurceller visar stora variationer i protein- och enzyminnehållet. Några av enzymerna är ADPas, ATPas, NADPH cytokrom-C-reduktas, glykosyltransferaser, galaktosyltransferas, tiaminpyrofosfat etc.
3. Kolhydrater :
Både växt- och djurceller har några vanliga kolhydratkomponenter, som glukosarin, galaktos, glukos, mannos och fruktos. Växten Golgi saknar sialinsyra, men det förekommer i hög koncentration i råttlever. Vissa kolhydrater som xylulos och arabinos är endast närvarande i växtceller.
4. Vitamin С :
Fraktionen av vitamin С som lagrats i Golgi-komplexet har visats av Tomitte. Enligt honom lagrar Golgi-komplexet vitamin С och frigör det långsamt i cytoplasman i tillräcklig mängd för att förhindra oxidation av cellprodukterna.

Golgikomplexets funktioner:
1. Formation av akrosom under spermiogenesen :
Under mognad av spermier spelar Golgi-komplexet en roll vid bildandet av akrosom (Burgos och Fawcett, 1955).

I tidiga skeden uppträder Golgi som en sfärisk kropp, bestående av cisternae arrangerad i parallella staplar och många små vesiklar. Den senare klämde alltid från cisternae. När utvecklingen fortsätter blir Golgi-komplexet oregelbundet i form och stora vakuloler bildas av dilateringar av cisternal sacs.
I mitten av dessa stora vakuoler eller vakuoler finns en tät granul, den proacrosomala granulen. Denna granul som härrör från Golgi-komplexet fortsätter växa i vakuolen med en process som kallas accretion. Denna vakuol och granulär närmar sig den främre polen av kärnmembranet, som utgör acrosomal granulat.
Med förlängningen av spermatiden sprider den acrosomala vesikelen över kärnans yta och sammanfaller slutligen med kärnmembranet och bildar lockmaterialet. Den acrosomala granulen blir det akrosom som ligger vid kärnans topp och innefattar uppenbarligen vissa enzymer som är involverade i fertiliseringsprocessen.
2. Syntes och utsöndring av polysackarider:
Studier av bägge celler genom autoradiografi och elektronmikroskopi har fastställt interrelationen mellan proteinsyntes, tillsats av kolhydrater och sulfatering. Colonens bägge celler producerar mucigen. Detta sekretoriska material innehåller en stor andel kolhydrat.
Golgi-komplexet ligger strax ovanför kärnan. Mot cellens fria yta förstoras gradvis mucigengranuler. Golgi-komplexets proximala cisternae visar ingen svullnad, men på något avstånd över stapeln omvandlas distala cisternae helt plötsligt till mucigengranuler. Den distala cisternaen omvandlas kontinuerligt till mucigengranulat varannan 2-4 minuter. Ny proximal cisternae bildas i ersättning.
3. Roll i sekretion:
Golgi-komplexet anses spela en roll i sekretorisk funktion hos en cell. Men frågan är att de utsöndrar eller syntetiserar vissa ämnen själva eller de är helt enkelt ett butikshus där de sekretoriska produkterna som utsöndras någon annanstans i cellen helt enkelt lagras och koncentreras.


Från studierna av Palade et al. 1962 är denna sekretoriska cykel nu väldefinierad och innehåller fyra steg vid akutceller i bukspottskörteln och de är:
(i) Införlivandet av aminosyror i protein vid ytan av grov endoplasmatisk retikulum.
(ii) Överföring av dessa växande sekretoriska proteiner i cisternaen av grov endoplasmisk retikulum.
(iii) Den intracellulära transporten av dessa proteiner till Golgi-komplexet.
(iv) Migrering av zymogengranuler mot cellens topp där de släpptes ut i lumen.
4. Golgi kroppens roll i oogenesen :
Srivastava (1965) har gett en kort genomgång av Golgi-komplexet under oogenes. Enligt Afzelius (1956) består Golgi-komplexet av ett hav-äggborrägg, som ses under elektronmikroskop, av staplar av lameller som bildar väggar av plana påsar, som ibland kan svullas.
Det finns några tecken på tvärgående uppdelningar av dessa kroppar. Sotelo (1959) och Sotelo och Porter (1959) har beskrivit Golgi-komplexet i råtta-ägg som ses under elektronmikroskopet och fann juxtra-kärnlokalisering av denna organell i tidiga oocyter.
I nästa steg löser de sig i fragment och i tredje etappen flyttas dessa mot cortexen. I alla dessa fall återstår deras struktur att vara av tätt packade arrays av smala dubbla profiler (plana sackar) och sfäriska vesiklar.
I tidiga oocyter är komplexet kompakt organiserat. I senare skeden finns diskreta buntar av profiler, omgivna av små vesiklar, utspridda i den kortikala zonen. I de tidiga oocyterna är Golgi-komplexet och centrosomen nära förbundna.
5. Absorption av föreningar :
Hirsch et al., Har upptäckt att när järn socker matas till ett djur, blir järnet absorberat på Golgi-komplexet (Kedrowsky). Van Teel har visat att Golgi system också absorberar föreningar av koppar och guld. Kedrowsky har visat att Golgi-komplexet av Opalina kan absorbera bismutos (förening av albumin och vismut) och protargol (förening av albumin och silver). Således anger Kirkman och Severinghaus att Golgi-komplexet fungerar som ett kondensationsmembran för koncentrationen av produkter i droppar eller granuler.
6. Växtcellsväggbildning :
Växtens cellvägg består av fibriller som övervägande innehåller polysackarider, tillsammans med vissa lipider och proteiner. Under cytokines bildas en cellplatta mellan de två dotterkärnorna och har därvid ett membran som senare blir dottercellernas plasmamembran. Det finns tydliga bevis för att polysackariderna bildas i Golgi-komplexet och överförs till den nya cellväggen som läggs ner medan cellerna fortfarande växer.

Ämnen som pektiner och hemicelluloser, som bildar matrisen av cellplattan som separerar plasmamembranen, bidrar också av Golgi-komplexet.
7. Formation av intracellulära kristaller :
I den marina isopoden är Limnoria lingmorum, som är en burrowing form, närvarande midglands, vars celler består av kristaller. Dessa intervall upp till 30 A ° i längd och 15 A ° tjocka. Det har bevisats att dessa kristaller bildas av Golgi-komplexet och är kända för att innehålla protein och järn. De är utan omslutande membran och vanligtvis sfäriska i form. De är oroade mot den sekretoriska verksamheten.
8. Droppbildning av mjölkprotein :
I den lakterande bröstkörteln hos möss produceras proteindroppar som är relaterade till Golgi-komplexet. Dessa droppar öppnar vanligtvis på cellytan genom fusion av deras inneslutande membran med plasmamembranet.
9. Bildning av lysosomer och vakuoler :
Primärlysosomer bildas från Golgi-membran på samma sätt som sekretoriska vesiklar. Det finns goda bevis på att diktyosomer ackumulerar hydrolytiska enzymer i sina mer mogna regioner. Vissa vacuoler i växtceller har visat sig innehålla små mängder hydrolytiska enzymer och dessa antas ha härlett från Golgi-komplexet.
10. Pigmentbildning:
I många däggdjurs- och cancerceller har Golgi-komplexet beskrivits som ursprungsorten för pigmentgranuler (melanin).
11. Reglering av vätskebalans :
En homologi har föreslagits mellan Golgi-komplexet och kontraktil vakuolen av lägre Metazoa och Protozoa. Kontraktil vacuolen utvisar överskott av vatten från cellen. I vissa Protozoer är Golgi-komplexet också oroad över reglering av vätskebalans.
Golgi-komplexets ursprung:
Tre olika källor har föreslagits från vilka nya Golgi-komplex kan uppstå:
1. Från endoplasmatisk retikulum :
Essner och Novik®ff (1962) och Beams and Kessel (1968) har föreslagit att Golgi-cisternaen kommer från ER. Den grova endoplasmatiska retikulum efter syntetisering av specifika proteiner förlorar ribosomer och förändras till jämn ER. Små övergående vesiklar kläms av från slät ER. Dessa migrerar till diktyosom.
När man når diktyosoms formningsyta bildas dessa säkringar för att bilda ny cisternae och därmed bidra till dess tillväxt. Genom sammansmältningen av dessa blåsor bildas nya cisternae kontinuerligt på formningsytan och på det mogna ansiktet bryter de gamla cisternaen ner i sekretoriska blåsor. Således uppvisar Golgi ett fenomen av membranflöde. '
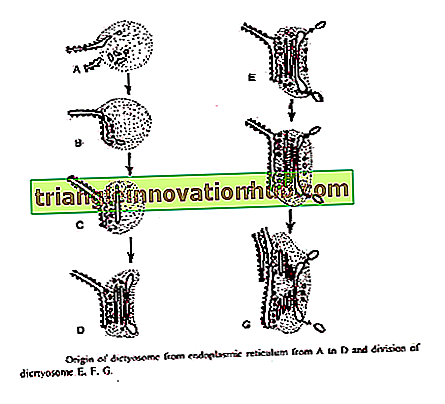

Bouch (1965) beskrev Golgis ursprung från yttermembran av kärnhölje i bruna alger. Vesiklarna klämmas av från yttre kärnmembran som smälter för att bilda cisternae på diktyosoms formningsyta.
Förekomsten av uteslutningszoner i samband med slät ER eller kärnmembran, förekomsten av uteslutningszoner i vilande frö från högre växter och bildandet av diktyosom från dessa zoner i groddfrön ger bevis till stöd för ovanstående teorier om ursprunget av dictyosome.
3. Genom uppdelning av existerande diktyosom :
Det har observerats att vid celldelning i både växter och djur ökar antalet diktyosomer och antalet diktyosomer i varje dottercell strax efter delningen är nästan lika med antalet i modercellen före delningen, från burkar och andra direkta observationer på delningscellerna har det antagits att diktyosomer också delar upp under celldelning.


