DNA-replikation: Anmärkningar om halv-konservativ replikering av DNA
Läs den här artikeln för att lära dig om DNA-replikationen: Anmärkningar om halv-konservativ replikering av DNA!
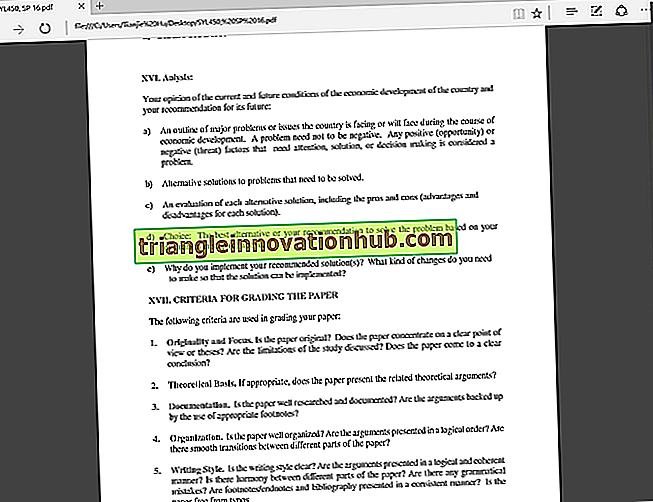
Replikering är processen för bildning av kolkopior. För detta fungerar DNA som sin egen mall. Därför är DNA-replikation en autokatalytisk funktion av DNA.

Image Courtesy: ehrig-privat.de/ueg/images/dna-replic.jpg
Det uppträder vanligtvis under S-fasen av cellcykeln när kromosomer är i mycket utsträckt form. Som föreslagits av Watson och Crick är DNA-replikation semikonservativ (en typ av replikering i vilken en sträng av dotterduplexen härrör från föräldern medan den andra strängen formas på nytt).
Detta utförs genom separation av två strängar. De separerade strängarna fungerar som mallar. De nya strängarna uppbyggda över mallarna av gamla strängar kommer att ha kompletterande baspar (ett motsatta T och G motsatt C). De två döttrarna DNA-molekyler som bildas på detta sätt kommer att vara kolkopior av modermolekylen men ska ha en ny sträng och en gammal sträng.
Taylor et al (1957) matade delande celler av rottips av Broad Bean (Vicia faba) med radioaktivt 3H innehållande tymin istället för normal tymin. Thymin införlivas i DNA som är det strukturella elementet i kromosomer. Taylor et al fann att alla kromosomer blev radioaktiva.
Märkt tymin ersattes sedan med normal en. Nästa generation kom till att ha radioaktivitet i en av de två kromatiderna av varje kromosom medan i efterföljande generationsradioaktivitet var närvarande i 50% av kromosomerna (fig 6.9). Detta är endast möjligt om en av de två strängarna av en kromosom bildas en gång i tiden medan den andra är konserverad vid varje replikation är detta semikonservativ replikation.

Halvkonservativ replikering av DNA bevisades av arbetet av Mathew Meselson och Franklin Stahl (1958). De växte Escherichia coli i många generationer i ett medium med tung isotop av kväve, i form av 15 NH4Cl tills bakteried DNA blev fullständigt märkt med tung isotop.
De märkta bakterierna skiftades sedan till färskt medium med normalt eller 14 N kväve. Prover togs för varje generation (en generation tar 20 minuter då E. coli delar upp på 20 minuter) och DNA testades för den tunga isotopen av kväve genom densitetsgradientcentrifugering med användning av cesiumklorid. Cesiumklorid är mycket vattenlösligt tungt salt.
När den centrifugeras i hög hastighet (säg 50 000 varv per minut) bildar saltet en densitetsgradient med den tyngsta mest koncentrerade regionen längst ner och successivt mindre koncentrerad lättare mot ytan. När DNA blandas med cesiumklorid kommer det att sätta sig ner vid en viss höjd i centrifugering, tyngre mot basen och ljusare en högre upp (Fig 6.10).
Fluorkrom som kallas etidiumbromid används för att förbättra kontrasten eftersom fluorokrom är specifik för DNA. Meselson och Stahl fann att DNA från den första generationen var hybrid eller intermediär ( 15 N och 14 N). Det avgjordes i cesiumkloridlösning vid en nivå högre än det fullständigt märkta DNA hos moderbakterierna ( 15 N 15 N). Den andra generationen bakterier efter 40 minuter innehöll två typer av DNA, 50% lätt ( 14 N I4 N) och 50% mellanprodukt ( 15 N I4 N).
Den tredje generationen bakterier efter 60 minuter innehöll två typer av DNA, 25% mellanprodukt ( 15 N 14 N) och 75% lätt ( 14 N 14 N) i 1: 3-förhållande. Den fjärde generationen efter 80 minuter innehöll 12, 5% 15 N 14 N och 87, 5% 14 N 14 N DNA i förhållandet 1: 7.
Denna observation är endast möjlig om de två strängarna av DNA-duplex separeras vid replikationstiden och fungerar som en mall för syntesen av nya komplementära strängar av DNA som har normal eller 14 N. Detta kommer att producera två DNA-duplexer med en gammal sträng ( 15 N) och en ny sträng ( 14 N).

Under bildandet av andra generationen separeras 15 N och 14 N strängar av DNA-duplex för att fungera som mallar så att 50% av nya DNA-duplexer endast har normala eller 14 N-strängar medan ytterligare 50% har både 15 N och 14 N-strängar (Figs 6, 11 och 6, 12). På detta sätt vid varje replikation bevaras en sträng av förälder-DNA i dottern medan den andra syntetiseras nyligen.